Генетическая инженерия
Современная биология коренным образом отличается от традиционной биологии не только большей глубиной разработки познавательных идей, но и более тесной связью с жизнью общества, с прак- тикой. Можно сказать, что в наше время биология стала средством преобразования живого мира с целью удовлетворения материальных потребностей общества. Это заключение иллюстрируется прежде всего тесной связью биологии с биотехнологией, которая стала важнейшей областью материального производства, равноправным партнером механической и химической технологий, созданных чело- веком, а также с медициной.
С момента своего возникновения биология и биотехнология всегда развивались совместно, причем с самого начала биология была научной основой биотехнологии. Однако длительное время недостаток собственных данных не позволял биологии оказывать очень большое влияние на биотехнологию. Положение резко изменилось с созданием во второй половине XX в. методологии генетической инженерии, под которой понимают генетическое манипулирование с целью конструкции новых и реконструкции существующих генотипов. Являясь по своей природе методическим достижением, генетическая инженерия не привела к ломке сложившихся представлений о биологических явлениях, не затронула основных положений биологии подобно тому, как радиоастрономия не поколебала основных положений астрофизики, установление «механического эквивалента тепла» не привело к изменению законов теплопроводности, а доказательство атомистической теории вещества не изме- нило соотношений термодинамики, гидродинамики и теории упругости (А.А. Баев).
Тем не менее генетическая инженерия открыла новую эру в биологии по той причине, что появились новые возможности для про- никновения в глубь биологических явлений с целью дальнейшей характеристики форм существования живой материи, более эффективного изучения структуры и функции генов на молекулярном уровне, понимания тонких механизмов работы генетического аппарата. Успехи генетической инженерии означают переворот в современном
естествознании. Они определяют критерии ценности современных представлений о структурно-функциональных особенностях молекулярного и клеточного уровней живой материи. Современные данные о живом имеют гигантское познавательное значение, ибо обеспечивают понимание одной из важнейших сторон органического мира и тем самым вносят неоценимый вклад в создание научной картины мира. Таким образом, резко расширив свою познавательную базу, биология через генетическую инженерию оказала также ведущее влияние на подъем биотехнологии.
Генетическая инженерия создает заделы на пути познания способов и путей «конструирования» новых или улучшения существующих организмов, придавая им большую хозяйственную ценность и способность резкого увеличения продуктивности биотехнологических процессов. Однако генетическая инженерия создала новые горизонты и для медицины по линии диагностики и лечения многих болезней, как ненаследственных, так и наследственных. Она открыла новые пути в поисках новых лекарств и материалов, используемых в медицине. Генетическая инженерия и биотехнология стимулировали разработку методов бионанотехнологии.
В рамках генетической инженерии различают генную и клеточную инженерию. Под генной инженерией понимают манипуляции с целью создания рекомбинантных молекул ДНК. Часто эту методологию называют молекулярным клонированием, клонированием генов, технологией рекомбинантных ДНК или просто генетическими манипуляциями. Важно подчеркнуть, что объектом генной инженерии являются молекулы ДНК, отдельные гены. Напротив, под клеточной инженерией понимают генетические манипуляции с изолированными отдельными клетками или группами клеток растений и животных.
ГЕННАЯ ИНЖЕНЕРИЯ И ЕЕ ИНСТРУМЕНТЫ
Генную инженерию составляет совокупность различных экспериментальных приемов (методик), обеспечивающих конструкцию (реконструкцию), клонирование молекул ДНК и генов с заданными целями.
Методы генной инженерии используют в определенной последовательности (рис. 127), причем различают несколько стадий в выпол-
нении типичного генно-инженерного эксперимента, направленного на клонирование какого-либо гена, а именно:
1. Выделение плазмидий ДНК из клеток интересующего организма (исходного) и выделение ДНК-вектора.
2. Разрезание (рестрикция) ДНК исходного организма на фрагменты, содержащие интересующие гены, с помощью одного из ферментов-рестриктаз и выделение этих генов из рестрикционной смеси. Одновременно разрезают (рестрицируют) векторную ДНК, превращая ее из кольцевой структуры в линейную.
3. Смыкание интересующего сегмента ДНК (гена) с ДНК вектора с целью получения гибридных молекул ДНК.
4. Введение рекомбинантных молекул ДНК путем трансформации в какой-либо другой организм, например в Е. coli или соматические клетки.
5. Высев бактерий, в которые вводили гибридные молекулы ДНК, на питательные среды, позволяющие рост только клеток, содержащих гибридные молекулы ДНК.
6. Идентификация колоний, состоящих из бактерий, содержащих гибридные молекулы ДНК.
7. Выделение клонированной ДНК (клонированных генов) и ее характеристика, включая секвентирование азотистых оснований в клонированном фрагменте ДНК.
Рис. 127. Последовательные стадии генно-инженерного эксперимента
В ходе эволюции бактерии развили способность синтезировать так называемые рестрицирующие ферменты (эндонуклеазы), которые стали частью клеточной (бактериальной) системы рестрикциимодификации. У бактерий системы рестрикции-модификации являются внутриклеточной иммунной системой защиты от чужеродной ДНК. В отличие от высших организмов, у которых распознание и разрушение вирусов, бактерий и других патогенов происходит внеклеточно, у бактерий защита от чужеродной ДНК (ДНК растений и животных, в организме которых они обитают) происходит внутриклеточно, т.е. тогда, когда чужеродная ДНК проникает в цитоплазму бактерий. С целью защиты бактерии в ходе эволюции развили также способность «метить» собственную ДНК метилирующими основаниями на определенных последовательностях. По этой же причине чужеродная ДНК из-за отсутствия в ней метильных групп на тех же последовательностях плавится (разрезается) на фрагменты разными бактериальными рестриктазами, а затем деградируется бактериальными экзонуклеазами до нулеотидов. Можно сказать, что таким образом бактерии защи- щают себя от ДНК растений и животных, в организме которых они обитают временно (как патогены) или постоянно (как сапрофиты).
Рестриктазы впервые были выделены из Е. coli в 1968 г. Оказалось, что они способны разрезать (плавить) молекулы ДНК на разных сайтах (местах) рестрикции. Эти ферменты получили название эндонуклеаз класса I. Затем у бактерий были обнаружены эндонуклеазы класса II, которые распознают в чужеродной ДНК сайты рестрикции специфически и на этих сайтах тоже осуществляют рестрикцию. Именно ферменты этого класса стали использовать в генной инже- нерии. Тогда же были открыты ферменты класса III, которые плавят ДНК рядом с сайтами распознания, но эти ферменты не имеют значения в генной инженерии.
Действие системы рестрикции-модификации «рационализуется» так называемыми палиндромными (распознающими) последователь- ностями азотистых оснований, которые являются сайтами рестрикции ДНК. Палиндромные последовательности - это последовательности оснований, которые одинаково читаются вперед и назад, как, например, последовательность букв радар. Поскольку цепи ДНК обладают антипараллельным направлением, то считают, что последовательность является палиндромной, если она идентична, когда читается в направлении от 5" - к 3"-концу на верхней и на нижней цепи от 3" - к 5"-концу, а именно:

Палиндромы могут быть любых размеров, но большинство тех палиндромов, которые используют в качестве сайтов узнавания рестриктазами, состоят из 4, 5, 6 и реже 8 оснований.
Рестриктазы - это абсолютно необходимый инструмент в генной инженерии для вырезания интересующих фрагментов (генов) из больших молекул ДНК. Поскольку известно более 100 ферментов рестрикции, то это позволяет выбор рестриктаз и селективное вырезание фрагментов из исходной ДНК.
Замечательной особенностью рестриктаз является то, что они продуцируют разрезы молекул на несколько фрагментов (рестриктов) ДНК уступами, в результате чего в образующихся концах одна цепь длиннее другой, образуя своеобразный хвост. Такие концы (хвосты) получили название «липких» концов, так как они способны к самокомплементарности.
Рассмотрим результаты рестрикции на примере одной из наиболее известных рестриктаз Eco RI из системы рестрикция-модификация Е. соИ. Вместо того чтобы плавить ДНК в центре палиндромной последовательности узнавания, этот фермент плавит ДНК за преде- лами центра и продуцирует 4 самокомплементарных («липких») конца, состоящих из разного количества нуклеотидов, а именно:

Эти «липкие» концы в генно-инженерных опытах полезны по той причине, что они могут быть воссоединены комплементарно при низких температурах, что позволяет эффективное смыкание ДНК-фрагментов.
Сайты распознавания и сайты плавления в случае других рестриктаз имеют другое содержание, а именно:

Вслед за рестрикцией ДНК из рестрикционной смеси выделяют рестрикционные ДНК-фрагменты (ДНК-рестрикты), которые необ- ходимы затем для объединения с вектором. Для выделения ДНКрестриктов прибегают к электрофорезу, поскольку с помощью этого метода рестрикцированную ДНК очень легко фракционировать благодаря размерам фрагментов-рестриктов и константным отношениям электрический заряд-масса. Фрагменты в электрическом поле мигрируют в ходе электрофореза при частоте, зависимой от их размеров (массы). Чем больше (длиннее) фрагмент, тем медленнее он мигрирует в электрическом поле. Материалом, в котором проводят электрофорез, являются незаряжающиеся агароза или полиакриламид. Для опознания фрагментов используют этидий бромид, который красит фрагменты, что ведет к их более легкому обнаружению.
Результативность электрофореза очень высока, поскольку с его помощью могут быть разделены фрагменты, размеры которых состав- ляют от 2 до 50 000 оснований.
После электрофореза фрагменты из агарозы выделяют с помощью разных методов. На основании результатов сравнения размеров
рестриктов одной и той же ДНК, полученных с помощью разных рестриктаз, строят рестрикционные карты, на которых показывают сайты рестрикции каждой из использованных рестриктаз. В практическом плане рестрикционные карты позволяют определять не только размеры рестриктов, но и выяснять расположение в молекулах ДНК локусов тех или иных генов.
Поскольку у высших организмов в ходе транскрипции синтезируется гетерогенная ДНК, корректируемая процессингом, то в генной инженерии обычно используют комплементарную ДНК (кДНК), которую получают при использовании в качестве матрицы мРНК, на которой обратная транскриптаза синтезирует одноцепочечную ДНК (кДНК), являющуюся копией мРНК. В последующем эти одноцепочечные ДНК превращают в двухцепочечные ДНК. Считают, что кДНК содержит непрерывные нуклеотидные последовательности (транскрибируемые и транслируемые). Именно кДНК используют для рестрикции.
Выделенные после электрофореза из агарозных гелей фрагменты ДНК (рестрикты) можно предварительно подвергнуть сек-вентированию, т.е. определить в них нуклеотидную последовательность. Для этого служат химический и ферментативный методы секвентирования. Химический метод основан на получении меченных радиоактивным фосфором (32 Р) фрагментов и удалении из этих фрагментов одного из оснований с последующим учетом результатов радиоавтографии гелей, содержащих эти фрагменты. Ферментативный метод основан на том, что в конец анализируемого фрагмента вводят нуклеотид, используемый затем в синтезе разных фрагментов in vitro, анализируемых на нуклеотидную последовательность электрофоретически. Для изучения специфических последовательностей нуклеотидов в молекуле ДНК используют
также гибридизацию ДНК-ДНК, РНК-РНК, ДНК-РНК, Нозерн-
и Саузерен-блоттинги.
Генетические векторы. Сегмент ДНК (ген), который предназначен для молекулярного клонирования, должен обладать способностью к репликации при переносе его в бактериальную клетку, т.е. быть репликоном. Однако он такой способностью не обладает. Поэтому, чтобы обеспечить перенос и обнаружение клонируемых генов в клетках, их объединяют с так называемыми генетическими векторами. Последние должны обладать как минимум двумя свойствами. Во-первых, векторы должны быть способны к репликации
в клетках, причем в нескольких концах. Во-вторых, они должны обеспечивать возможность селекции клеток, содержащих вектор, т.е. обладать маркером, на который можно вести контрселекцию клеток, содержащих вектор вместе с клонируемым геном (рекомбинантные молекулы ДНК). Таким требованиям отвечают плазмиды и фаги. Плазмиды являются хорошими векторами по той причине, что они являются репликонами и могут содержать гены резистентности к какому-либо антибиотику, что позволяет вести селекцию бактерий на устойчивость к этому антибиотику и, следовательно, легкое обнаружение рекомбинантных молекул ДНК
(рис. 128).
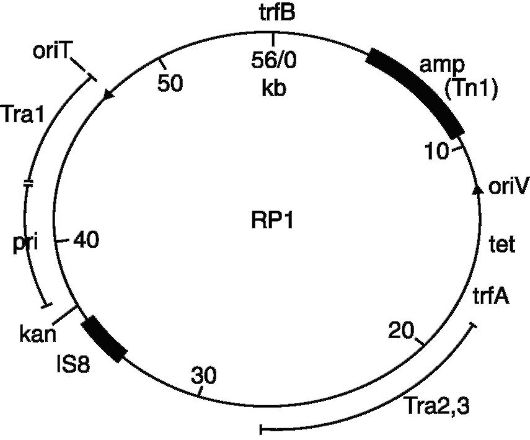
Рис. 128. Вектор pBRl
Поскольку не существует природных плазмидных векторов, то все известные к настоящему времени плазмидные векторы были сконструированы искусственно. Исходным материалом для создания ряда генетических векторов послужили R-плазмиды, в которых с помощью рестриктаз удаляли излишние последовательности ДНК, в том числе те, на которых располагались множественные сайты рестрикции. Это удаление определялось тем, что плазмидный вектор должен обладать только одним сайтом узнавания для одной рестриктазы, причем этот сайт должен лежать в функционально несущественном районе плазмидного генома. Например, плазмидый вектор pBR 322, который имеет гены резистентности к ампициллину и тетрациклину, что делает его очень удобным
для селекции бактерий, содержащих клонируемый сегмент ДНК, обладает одиночными сайтами рестрикции для более 20 ферментов- рестриктаз, включая такие известные рестриктазы, как Eco RI, Hind III, Pst I, Pva II и Sal I.
Фаговые векторы тоже обладают рядом преимуществ. Они могут включать в себя более крупные (более длинные) клонируемые фрагменты ДНК по сравнению с плазменными векторами. Далее, перенос фагами клонируемого фрагмента в клетки в результате инфицирования ими последних является более эффективным, чем трансформация ДНК. Наконец, фаговые векторы позволяют более эффективный скрининг (распознание) на поверхности агара колоний, содержащих клетки, несущие клонируемый ген. Многие фаговые векторы сконструированы на базе фага лямбда.
Кроме фаговых используют и другие вирусные векторы, сконструированные на базе вируса герпеса, а также векторы, сконструированные на базе дрожжевой ДНК.
Если клонирование генов проводят, используя клетки млекопитающих или растений, то требования к векторам те же, что и в случае клонирования в бактериальных клетках.
Конструирование рекомбинантных молекул ДНК. Непосредственное конструирование рекомбинантных молекул ДНК следует после того, как получены рестрикты исследуемой ДНК и векторной ДНК. Оно заключается в смыкании сегментов-рестриктов исследуемой ДНК с рестриктом векторной ДНК, которая в результате рестрикции превращается из кольцевой в линейную ДНК.
Чтобы сомкнуть фрагменты исследуемой ДНК с ДНК вектора, используют ДНК-лигазу (рис. 129). Лигирование будет успешным, если смыкаемые структуры обладают З"-гидроксильной и 5"-фос- фатной группами и если эти группы расположены соответствующим образом относительно одна другой. Фрагменты объединяются через их «липкие» концы в результате самокомплементарности. При высоких концентрациях фрагментов последние время от времени становятся в правильное положение (напротив друг друга). Многие рестриктазы, такие как Eco RI, продуцируют «липкие» концы, состоящие из четырех оснований. Процесс лигирования «липких» концов, состоящих из четырех оснований, происходит при пониженной температуре (до 12 ?С).
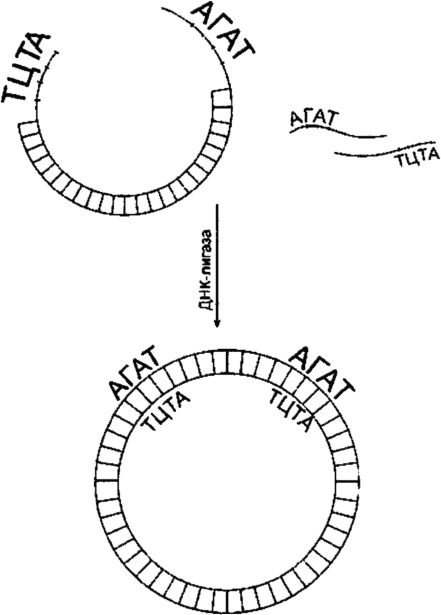
Рис. 129. ДНК-лигирование
Если при рестрикции образуются фрагменты без «липких» концов, то их «насильственно» конвертируют в молекулы с «липкими» концами, используя фермент трансферазу. Этот фермент добавляет нуклеотиды к 3"-концу ДНК. На одном фрагменте может быть добавлен поли-А-хвост, на другом - поли-Т-хвост. Для генерации любых желаемых концов ДНК используют также полимеразную цепную реакцию (ПЦР). Принцип ПЦР основан на денатурации выделенной из клеток ДНК и «отжиге» ее с добавлением к ренатурирующимся цепям ДНК-олигонуклеотидов, состоящих из 15-20 нуклеотидов каждый. Эти олигонуклеотиды должны быть комплементарны последовательностям в цепях, разделенных расстояниями в 50-2000 нуклеотидов. Будучи «затравкой» для синтеза ДНК in vitro, они позволяют ДНК-полимеразе копировать те участки, которые находятся между «затравками». Это копирование дает большое количество копий изучаемого фрагмента ДНК.
Введение рекомбинантных молекул ДНК в клетки. После смыкания интересующего фрагмента ДНК (гена) с генетическим вектором с помощью ДНК-лигазы образованные рекомбинантные молекулы вводят в клетки с целью добиться их репликации (за счет генетического вектора) и увеличения количества копий. Наиболее популярным способом введения в клетки рекомбинантных молекул ДНК, в которых вектором служит плазмида, является трансформация Е. coli. С этой целью бактериальные клетки предварительно обрабатывают кальцием или рубидием (ионами), для того
чтобы они стали «компетентными» в восприятии рекомбинатной ДНК. Чтобы повысить частоту проникновения ДНК в клетки, используют метод электропорации, заключающийся в кратком экспонировании клеток в интенсивном электрическом поле. Эта обработка создает полости в мембранах клеток, что способствует лучшему восприятию клетками ДНК. После введения рекомбинатных молекул ДНК в бактерии последние высевают на МПА (мясо-пептонный агар), обогащенный антибиотиками для селекции желаемых клеток, т.е. клеток, содержащих рекомбинантные молекулы ДНК. Частота трансформации является невысокой. Обычно один трансформант возникает на 10 5 высеянных клеток. Если же вектор является фаговым, то прибегают к трансфекции клеток (бактерий или дрожжей) фагом. Что касается соматических клеток животных, то их трансфекцию осуществляют ДНК в присутствии химических веществ, облегчающих прохождение ДНК через плазматические мембраны. Возможны также прямые микроинъекции ДНК в овоциты, в культивируемые соматические клетки и в эмбрионы млекопитающих.
Важнейшим моментом, связанным с молекулярным клонированием, является поиск способа, позволяющего установить, действитель- но ли клонируемый фрагмент включился в вектор и вместе с вектором, образовав рекомбинатную молекулу ДНК, вошел в клетки. Если речь идет о бактериальных клетках, то один из способов основан на учете инсерционной инактивации плазмидного (векторного) гена резистентности. Например, в плазмидном векторе pBR 322, детерминирующем резистентность к ампициллину и тетрациклину, един- ственный сайт для рестриктазы Pst I находится в локусе, занимаемом геном резистентности к ампициллину. Pst I-плавление на этом сайте генерирует «липкие» концы, позволяющие лигирование клонируемого фрагмента с векторной ДНК. Однако при этом плазмидный (векторный) ген ампициллинрезистентности инактивируется, тогда как ген тетрациклинрезистентности на векторе остается интактным. Именно ген тетрациклинрезистентности и используется для селекции клеток, трансформируемых рекомбинантными молекулами ДНК. Это позволяет убедиться, что клетки выросших колоний на среде с тетрациклином действительно содержат рекомбинантные молекулы ДНК, их проверяют с помощью так называемого «спот-теста» на паре чашек с плотной средой, одна из которых содержит ампициллин, тогда как другая лишена этого антибиотика. Клонируемые ДНК находятся
лишь в трансформантах, резистентных к тетрациклину. Что касается трансформантов, резистентных одновременно к ампициллину и тетрациклину (АрТс), то они содержат плазмидные (векторные) молекулы, которые спонтанно приобрели кольцевую форму без включения в них чужеродной (клонируемой) ДНК.
Другой способ обнаружения инсерции чужеродных (клонируемых) фрагментов в плазмидный вектор основан на использовании вектора, содержащего ген β-галактозидазы. Инсерция чужеродной ДНК в этот ген неизбежно инактивирует синтез β-галактозидазы, что может быть обнаружено посевом трансформированных клеток на среду, которая содержит субстраты β-галактозидазы. Эта среда позволяет селекцию окрашенных колоний клеток. Существуют и другие методы.
Как уже отмечено, рестрикционные линейные фрагменты векторной ДНК способны к восстановлению кольцевой структуры без включения в них клонируемых сегментов. Чтобы уменьшить частоту спонтанного образования таких кольцевых молекул векторной ДНК, рестрикты векторной ДНК обрабатывают фосфатазой. В результате этого образование кольцевых молекул ДНК становится невозможным, поскольку будут отсутствовать концы 5"-РО 4 , необходимые для действия лигазы.
Совокупность колоний-трансформантов, выросших на селективной среде, представляет собой совокупность клеток, содержащих клоны разных фрагментов (генов) клонируемой геномной или кДНК. Коллекции этих клонов формируют так называемые библиотеки ДНК, широко используемые в генно-инженерных работах.
Заключительной стадией клонирования генов является выделение и исследование клонированной ДНК, включая секвенирование. Перспективные штаммы бактерий или соматических клеток, содержащих рекомбинантные молекулы ДНК, которые контролируют синтез интересующих белков, имеющих коммерческую ценность, передают в промышленность.
КЛЕТОЧНАЯ ИНЖЕНЕРИЯ
Как отмечено в начале главы, клеточной инженерией называют генетические манипуляции с изолированными клетками животных и растений. Эти манипуляции часто осуществляют in vitro, а главной целью они имеют получение генотипов этих организмов с заданными свойствами, в первую очередь хозяйственно полезными. Что касает-
ся человека, то клеточная инженерия оказалась применимой к его половым клеткам.
Предпосылкой к развитию клеточной инженерии у человека и животных явилась разработка методов культивирования их сома- тических клеток на искусственных питательных средах, а также получение гибридов соматических клеток, включая межвидовые гибриды. В свою очередь успехи в культивировании соматических клеток оказали влияние на изучение половых клеток и оплодотворения у человека и животных. Начиная с 60-х гг. ХХ в. в нескольких лабораториях мира были выполнены многочисленные эксперименты по пересадке ядер соматических клеток в яйцеклетки, искусственно лишенные ядер. Результаты этих экспериментов часто были противоречивы, но в целом они привели к открытию способности клеточных ядер обеспечивать нормальное развитие яйцеклетки (см. гл. IV).
На основе результатов изучения развития оплодотворенных яйцеклеток в 60-е гг. XX в. были начаты также исследования по выяснению возможности оплодотворения яйцеклеток вне организма матери. Очень быстро эти исследования привели к открытию возможности оплодотворения яйцеклеток сперматозоидами в пробирке и дальнейшего развития образованных таким путем зародышей при имплантации в матку женщины. Дальнейшее совершенствование разработанных в этой области методов привело к тому, что рождение «пробирочных» детей стало реальностью. Уже к 1981 г. в мире было рождено 12 детей, жизнь которым была дана в лаборатории, в пробирке. В настоящее время этот раздел клеточной инженерии получил большое распространение, а количество «пробирочных» детей составляет уже десятки тысяч (рис. 130). В России работы по получению «пробирочных» детей были начаты только в 1986 г.
В 1993 г. была разработана методика получения монозиготных близнецов человека in vitro путем разделения эмбрионов на бласто- меры и доращивания последних до 32 клеток, после чего они могли быть имплантированы в матку женщины.
Под влиянием результатов, связанных с получением «пробирочных» детей, у животных тоже была разработана технология, получившая название трансплантации эмбрионов. Она связана с разработкой способа индукции полиовуляции, способов искусственного оплодот- ворения яйцеклеток и имплантации зародышей в организм животных - приемных матерей. Суть этой технологии сводится к следую-
щему. Высокопродуктивной корове вводят гормоны, в результате чего наступает полиовуляция, заключающаяся в созревании сразу 10-20 клеток. Затем яйцеклетки искусственно оплодотворяются мужскими половыми клетками в яйцеводе. На 7-8-й день зародыши вымывают из матки и трансплантируют в матки другим коровам (приемным матерям), которые затем дают жизнь телятам-близнецам. Телята наследуют генетический статус своих подлинных родителей.

Рис. 130. «Пробирочные» дети
Другой областью клеточной инженерии у животных является создание трансгенных животных. Наиболее простой способ получения таких животных заключается во введении в яйцеклетки исходных животных линейных молекул ДНК. Животные, развившиеся из оплодотворенных таким образом яйцеклеток, будут содержать в одной из своих хромосом копию введенного гена и, кроме того, они будут передавать этот ген по наследству. Более сложный способ получения трансгенных животных разработан на мышах, различающихся по окраске шерстного покрова, и сводится к следующему. Вначале из организма беременной серой мыши извлекают четырехдневных зародышей и измельчают их на отдельные клетки. Затем из эмбриональных клеток извлекают ядра, переносят их в яйцеклетки черных мышей, предварительно лишенных ядер. Яйцеклетки черных мышей, содержащие чужие ядра, помещают в пробирки
с питательным раствором для дальнейшего развития. Развившиеся из яйцеклетки черных мышей зародыши имплантируют в матки белых мышей. Таким образом, в этих экспериментах удалось получить клон мышей с серой окраской шерстного покрова, т.е. клонировать эмбриональные клетки с заданными свойствами. В главе IV мы рассмотрели результаты оплодотворения искусственно лишенных ядер яйцеклеток овец ядерным материалом соматических клеток животных этого же вида. В частности, из яйцеклеток овец удаляли ядра, а затем в такие яйцеклетки вводили ядра соматических клеток (эмбриональных, плодовых или клеток взрослых животных), после чего оплодотворенные таким образом яйцеклетки вводили в матки взрослых овец. Рождающиеся ягнята оказались идентичными овцедонору. Пример - овца Долли. Получены также клоновые телята, мыши, кролики, кошки, мулы и другие животные. Такое конструирование трансгенных животных представляет собой прямой путь клонирования животных с хозяйственно полезными признаками, включая особей определенного пола.
Трансгенные животные получены также при использовании исходного материала, принадлежащего разным видам. В частности, известен способ передачи гена, контролирующего гормон роста, от крыс в яйцеклетки мышей, а также способ комбинирования бластомеров овцы с бластомерами козы, что привело к возникновению гибридных животных (ковец). Эти эксперименты указывают на воз- можность преодоления видовой несовместимости на самых ранних этапах развития. Особенно заманчивые перспективы открываются (если видовая несовместимость будет преодолена полностью) на пути оплодотворения яйцеклеток одного вида ядрами соматических клеток другого вида. Речь идет о реальной перспективе создания хозяйственно ценных гибридов животных, которых невозможно получить путем скрещиваний.
Следует отметить, что ядерно-трансплантационные работы еще не очень эффективны. Эксперименты, выполненные на земноводных и млекопитающих, в целом показали, что их результативность является небольшой, причем она зависит от несовместимости между донорскими ядрами и реципиентными овоцитами. Кроме того, препятствием на пути к успехам являются также образующиеся хромосомные аберрации в трансплантированных ядрах в ходе даль- нейшего развития, которые сопровождаются гибелью трансгенных животных.
На стыке работ по изучению гибридизации клеток и иммунологических исследований возникла проблема, связанная с получением и изучением так называемых моноклональных антител. Как отмечено выше, антитела, продуцируемые организмом в ответ на введение антигена (бактерии, вирусы, эритроциты и т.д.), представляют собой белки, называемые иммуноглобулинами и составляющие фундаментальную часть защитной системы организма против возбудителей болезней. Но любое чужеродное тело, вводимое в организм, представляет собой смесь разных антигенов, которые будут возбуждать продукцию разных антител. Например, эритроциты человека обладают антигенами не только для групп крови А (II) и В (III), но и многими другими антигенами, включая резус-фактор. Далее, белки клеточной стенки бактерий или капсида вирусов также могут действовать в качестве разных антигенов, вызывающих образование разных антител. В то же время лимфоидные клетки иммунной системы организма обычно представлены клонами. Значит, даже только по этой причине в сыворотке крови иммунизированных животных антитела всегда представляют собой смесь, состоящую из антител, продуцируемых клетками разных клонов. Между тем для практических потребностей необходимы антитела только одного типа, т.е. так называемые моноспецифические сыворотки, содержащие антитела только одного типа или, как их называют, моноклональные антитела.
В поисках методов получения моноклональных антител швейцарскими исследователями в 1975 г. был открыт способ гибридизации между лимфоцитами мышей, иммунизированных тем или иным антигеном, и культивируемыми опухолевыми клетками костного мозга. Такие гибриды получили название «гибри- домные». От «лимфоцитарной» части, представленной лимфоцитом одного клона, одиночная гибридома наследует способность вызывать образование необходимых антител, причем одного типа, а благодаря «опухолевой (миэломной)» части она становится способной, как и все опухолевые клетки, бесконечно долго размно- жаться на искусственных питательных средах, давая многочисленную популяцию гибридов. На рис. 131 показана схема выделения клеточных линий, синтезирующих моноклональные антитела. Линии мышиных клеток, синтезирующих моноклональные антитела, выделяют путем слияния миеломных клеток с лимфоцитами из селезенки мыши, иммунизированной за пять дней до этого
желаемым антигеном. Слияния клеток достигают смешиванием их в присутствии полиэтиленгликоля, который индуцирует слияние клеточных мембран, а затем в высеве их на питательную среду, позволяющую рост и размножение только гибридных клеток (гибридом). Размножение гибридом проводят в жидкой среде, где они растут далее и секретируют антитела в культуральную жидкость, причем только одного типа, к тому же в неограниченных количествах. Эти антитела получили название моноклональных. Чтобы повысить частоту образования антител, прибегают к клонированию гибридом, т.е. к селекции отдельных колоний гибридом, способных вызывать образование наибольшего количе- ства антител желаемого типа. Моноклональные антитела нашли широкое применение в медицине для диагностики и лечения ряда болезней. В то же время важнейшее преимущество моноклональной технологии заключается в том, что с ее помощью могут быть получены антитела против материалов, которые невозможно очистить. Напротив, можно получить моноклональные антитела против клеточных (плазматических) мембран нейронов животных. Для этого мышей иммунизируют выделенными мембранами нейронов, после чего их селезеночные лимфоциты объединяют с миеломными клетками, а дальше поступают, как описано выше.

Рис. 131. Получение моноклональных антител
ГЕНЕТИЧЕСКАЯ ИНЖЕНЕРИЯ И МЕДИЦИНА
Генная инженерия оказалась очень перспективной для медицины, прежде всего в создании новых технологий получения физиологически активных белков, используемых в качестве лекарств (инсулин, соматостатин, интерфероны, соматотропин и др.).
Инсулин используют для лечения больных диабетом, который стоит на третьем месте (после болезней сердца и рака) по частоте вызываемых смертельных случаев. Мировая потребность инсулина составляет несколько десятков килограммов. Традиционно его получают из панкреатических желез свиней и коров, но гормоны этих животных слегка отличаются от инсулина человека. Инсулин свиней различается по одной аминокислоте, а коровий - по трем. Считают, что инсулин животных часто вызывает побочные эффекты. Хотя химический синтез инсулина осуществлен давно, но до сих пор промышленное производство гормонов оставалось очень дорогим. Сейчас получают дешевый инсулин с помощью генно-инженерного метода путем химико-ферментативного синтеза гена инсулина с последующим введением этого гена в кишечную палочку, которая затем синтезирует гормон. Такой инсулин более «биологичен», так как химически идентичен инсулину, вырабатываемому клетками поджелудочной железы человека.
Интерфероны - белки, синтезируемые клетками главным образом в ответ на заражение организма вирусами. Интерфероны характери- зуются видовой специфичностью. Например, у человека установлены три группы интерферонов, продуцируемых различными клетками под контролем соответствующих генов. Интерес к интерферонам определяется тем, что их широко используют в клинической практике для лечения многих болезней человека, особенно вирусных.
Имея крупные размеры, молекулы интерферона мало доступны для синтеза. Поэтому большинство интерферонов сейчас получают из крови человека, но выход при таком способе получения небольшой. Между тем потребности в интерфероне исключительно велики. Это поставило задачу изыскать эффективный метод производства интерферона в промышленных количествах. Генетическая инженерия лежит в основе современного производства «бактериального» интерферона.
Усилилось влияние генетической инженерии на технологию тех лекарственных веществ, которые уже давно создаются по био- логической технологии. Еще в 40-50-е гг. XX в. была создана
биологическая промышленность для производства антибиотиков, которые составляют наиболее эффективную часть лекарственного арсенала современной медицины. Однако в последние годы отмечается значительный рост лекарственной устойчивости бактерий, особенно к антибиотикам. Причина заключается в широком распространении в микробном мире плазмид, детерминирующих лекарственную устойчивость бактерий. Именно поэтому многие знаменитые ранее антибиотики утратили свою былую эффективность. Единственный пока путь преодоления резистентности бактерий к антибиотикам - это поиски новых антибиотиков. По оценкам специалистов, в мире ежегодно создают около 300 новых антибиотиков. Однако большинство из них либо неэффективно, либо токсично. В практику же каждый год вводится лишь несколько антибиотиков, что заставляет не только сохранять, но и увеличивать мощность антибиотической промышленности на основе генно-инженерных разработок.
Основные задачи генной инженерии в тех технологиях лекарственных веществ, в которых продуцентами лекарств являются микроорганизмы, определяются необходимостью генно-инженерной реконструкции последних с целью повышения их активности. В то же
время началась реализация идеи создания лекарств в виде малых молекул, что способствует их большей эффективности.
Иммунная биотехнология связана с производством прежде всего вакцин нового поколения для профилактики инфекционных болезней человека и животных. Первыми коммерческими продуктами, созданными с помощью генетической инженерии, стали вакцины против гепатита людей, ящура животных и некоторые другие. Исключительно важное направление в этой области связано с производством моноклональных антител, реагентов, необходимых для диа- гностики возбудителей болезни, а также для очистки гормонов, витаминов, белков различной природы (ферментов, токсинов и др.).
Значительный практический интерес представляет метод получения искусственного гемоглобина путем введения гемоглобиновых генов в растения табака, где под контролем этих генов продуциру- ются α- и β-цепи глобина, которые объединяются в гемоглобин. Синтезируемый в клетках табачных растений гемоглобин полностью функционален (связывает кислород). Клеточная инженерия в применении к человеку связана не только с решением фундаментальных проблем биологии человека, но и с преодолением прежде всего женского бесплодия. Поскольку частота положительных случаев имплантации в матку женщин эмбрионов, полученных in vitro, является небольшой, то получение монозиготных близнецовэмбрионов in vitro также имеет значение, так как увеличиваются возможности повторных имплантаций за счет «запасных» эмбрионов. Особый интерес представляют перспективы использования стволовых клеток в качестве источника замены клеток и тканей в лечении таких болезней, как диабет, повреждения спинного мозга, боли сердца, остиоартриты, болезнь Паркинсона. Но для реализации этих перспектив необходимо углубленное изучение биологии стволовых клеток.
В использовании генетической инженерии применительно к проблемам медицины особое значение приобрела задача разработки генно-инженерных методов радикального лечения наследственных болезней, которые, к сожалению, еще не поддаются лечению существующими методами. Содержание этой задачи заключается в разработке способов исправления (нормализации) мутаций, результатом которых являются наследственные болезни, и в обеспечении передачи «исправлений» по наследству. Считают, что успешной разработке генно-инженерных методов лечения наследственных болезней будут
способствовать данные о геноме человека, полученные в результате выполнения международной научной программы «Геном человека».
ЭКОЛОГИЧЕСКИЕ ПРОБЛЕМЫ ГЕНЕТИЧЕСКОЙ ИНЖЕНЕРИИ
Подняв на новый уровень биотехнологию, генетическая инженерия нашла также применение в разработке способов определения и устранения загрязнений окружающей среды. В частности, сконструированы штаммы бактерий, которые являются своеобразными индикаторами мутагенной активности химических загрязнений. С другой стороны, генно-инженерным способом сконструированы штаммы бактерий, содержащие плазмиды, под контролем которых происходит синтез ферментов, способных разрушать многие химические соединения- загрязнители среды обитания. В частности, некоторые плазмидосодержащие бактерии способны разлагать до безвредных соединений нефть и нефтепродукты, оказавшиеся в среде в результате различных аварий или других неблагоприятных причин.
Однако генетическая инженерия - это превращение генетического материала, которое в природе отсутствует. Следовательно, продукты генной инженерии - это абсолютно новые продукты, не существующие в природе. Поэтому она сама по себе из-за неизвестности ее продуктов таит опасность как для природы и среды обитания, так и для персонала, работающего в лабораториях, где используют методы генетической инженерии или работают со структурами, созданными в ходе генно-инженерных работ.
Поскольку возможности клонирования генов безграничны, то еще в самом начале этих исследований среди ученых возникли вопросы о природе создаваемых организмов. Одновременно были высказаны предположения о ряде нежелательных последствий этой методологии, причем эти предположения нашли поддержку и среди широкой общественности. В частности, появились раз- ногласия о свойствах бактерий, получивших в генно-инженерных экспериментах гены животных. Например, сохраняют ли бактерии Е. coli свою видовую принадлежность из-за содержания введенных в них генов животного происхождения (например, гена инсулина) или их следует считать новым видом? Далее, насколько долговечны такие бактерии, в каких экологических нишах они могут
существовать? Но самое главное стало заключаться в появлении опасений, что в ходе производства и манипуляций с рекомбинантными молекулами ДНК могут быть созданы генетические структуры со свойствами, непредвиденными и опасными для здоровья человека, для исторически сложившегося экологического равновесия. Тогда же начались и призывы к мораторию на генетическую инженерию. Эти призывы вызвали международный резонанс и привели к международной конференции, которая состоялась в 1975 г. в США и на которой широко обсуждались возможные последствия исследований в этой области. Затем в странах, где стала развиваться генетическая инженерия, были выработаны правила работы с рекомбинантными молекулами ДНК. Эти правила направлены на исключение попадания в среду обитания продуктов деятельности генно-инженерных лабораторий.
Другой аспект нежелательных последствий генно-инженерных работ связан с опасностью для здоровья персонала, работающего в лабораториях, где применяют методы генетической инженерии, поскольку в таких лабораториях используют фенол, этидий бромид, УФ-излучения, которые являются вредными для здоровья факторами. Кроме того, в этих лабораториях существует возможность заражения бактериями, содержащими рекомбинантные молекулы ДНК, контролирующие нежелательные свойства, например лекарственную резистентность бактерий. Эти и другие моменты определяют необходимость повышения уровня техники безопасности в генноинженерных работах.
Наконец, широко обсуждаются в обществе проблемы опасности генетически модифицированных продуктов (генетически изме- ненных томатов, картофеля, кукурузы, сои), а также таких продуктов, как хлеб, пасты, конфеты, мороженое, сыр, растительное масло, мясные продукты, которые в ряде стран, особенно в США, приобрели широкое распространение. На протяжении 12 000 лет сельского хозяйства человек употреблял продукты естественного происхождения. Поэтому предполагают, что с генетически модифицированной пищей в организм человека попадут новые токсины, аллергены, бактерии, канцерогены, что приведет к совершенно новым болезням будущих поколений. В связи с этим возникает вопрос о подлинно научной оценке генетически модифицированной пищи.
ВОПРОСЫ ДЛЯ ОБСУЖДЕНИЯ
1. Что понимают под генной, клеточной и генетической инженерией? Есть ли разница между этими понятиями и молекулярном клонировании?
2. В чем заключается прогрессивность генетической инженерии по сравнению с другими методами, используемыми в биологии?
3. Перечислите основные «инструменты» генной инженерии.
4. Что представляют собой ферменты-рестриктазы, каковы их свойства и их роль в генной инженерии?
5. Все ли рестриктазы образуют «липкие» концы исследуемых ДНК и зависит ли структура «липких» концов от вида рестриктазы?
6. Дайте определение генетическим векторам. Существуют ли природные векторы?
7. Как получают генетические векторы в лабораторных условиях? Какие биологические объекты являются исходным материалом для получения векторов?
8. Какова предельная длина последовательностей азотистых оснований ДНК, которые еще могут включиться в генетический вектор? Различаются ли векторы по «мощности»?
9. Охарактеризуйте свойства ДНК-лигазы и определите ее роль в генной инженерии.
10. Как смыкают клонируемый сегмент ДНК (ген) с генетическим вектором?
11. Какова частота введения рекомбинантных молекул ДНК в бактериальные клетки?
12. На каком принципе основана селекция бактериальных клеток, содержащих рекомбинантные молекулы ДНК? Приведите один из примеров такой селекции.
14. Многие штаммы бактерий обладают одинаковыми ферментами, практически одинаково обеспечивающими их метаболизм. Между тем нуклеотидная специфичность систем рестрикции-модификации бактерий различна. Можете ли вы объяснить это явление?
15. Почему последовательности ДНК, представляющие сайты распознавания рестриктазами, не могут содержать более восьми пар оснований?
16. Сколько раз последовательность ГГЦЦ, распознаваемая рестриктазой Нае III, будет встречаться в сегменте ДНК длиной в 50 000 пар оснований с 30-, 50- и 70-процентным содержанием ГЦ?
17. Рестриктазы Bam HI и Bgl I плавят последовательности Г ГАТЦЦ и Т ГАТЦА соответственно. Можно ли включить в сайт Bam HI фрагменты ДНК, продуцированные Bgl I-рестрикцией? Если да, то почему? Если используемая плазмида (вектор) содержит один сайт для рестрикции Bgl I, то на какой питательной среде можно осуществить селекцию бактерий, эту плазмиду?
18. Вычислите частоту трансформации бактерий на одну молекулу ДНК, если на 5000 плазмидных пар оснований образуется 5-10 5 трансформантов?
19. Можно ли клонировать 0-пункт репликации ДНК Е. coli и если да, то каким образом?
20. Можно ли определить, сколько необходимо молекул ДНК для трансформации одной клетки Е. coli?
21. Можно ли с помощью полимеразной цепной реакции определить сайт сплайсинга на мРНК?
22. Каким образом можно использовать полимеразную цепную реакцию для того, чтобы ввести желаемый сайт рестрикции в интересующее место на фрагменте ДНК, предназначенном для клонирования?
23. Назовите методы клеточной инженерии в применении к животным. Какова хозяйственная ценность животных, получаемых этими методами?
24. Дайте определение понятиям «трансгенные растения» и «трансгенные животные». Сохраняют ли трансгенные организмы свою видовую принадлежность или их можно считать организмами новых видов?
25. Что такое гибридомы и моноклональные антитела? Как их получают?
26. Применима ли клеточная инженерия к человеку?
27. Допустим, что инъекция чужеродной ДНК в яйцеклетку мыши и имплантация оплодотворенной таким путем яйцеклетки в организм мыши закончились ее беременностью и рождением мышат, содержащих в геноме копии инъецированной ДНК. Однако мышата оказались мозаиками, т.е. одни их клетки содержат копии инъецированной ДНК, другие лишены этой ДНК. Можете ли вы объяснить природу этого явления?
28. Считаете ли вы генетически опасной пищу, приготовленную из генетически измененных продуктов?
29. Необходима ли научная экспертиза генетически измененных продуктов питания?
Познание определяется тем, что утверждается нами как Истина.
П.А. Флоренский, 1923
Генная инженерия является совокупностью способов, приемов и технологий выделения из клеток или организма генов, получения рекомбинантных РНК и ДНК, осуществления различных манипуляций с генами, а также введения их и в другие организмы. Эта дисциплина способствует получению желаемых характеристик изменяемого организма.
Наукой в широком смысле генная инженерия не является, однако считается биотехнологическим инструментом. В ней применяются исследования таких наук, как генетика, молекулярная микробиология.
Созданные методы генной инженерии, связанные с управлением наследственностью, явились одним из самых ярких событий в развитии науки.
Ученые, молекулярные биологи, биохимики научились изменять, модифицировать гены и создавать абсолютно новые, сочетая гены от разных организмов. Также они научились и синтезировать материал в соответствии с заданными схемами. Искусственный материал ученые начали вводить в организмы, заставляя их работать. На всей этой работе основывается генная инженерия.
Однако существует некоторая ограниченность «биологического материала». Данную проблему ученые пытаются решить при помощи и Специалисты отмечают, что этот путь достаточно перспективен. В течение последних нескольких десятилетий учеными были созданы приемы, при помощи которых определенные клетки из растительных или можно заставить развиваться и размножаться независимо, отдельно от организма.
Достижения генной инженерии имеют большое значение. применяются в экспериментах, а также при промышленном производстве определенных веществ, которые невозможно получить при использовании бактериальных культур. Однако и в этой области имеют место трудности. Так, например, проблемой является отсутствие способности у клеток животных делиться такое же бесконечное количество раз, как
В ходе экспериментов были сделаны фундаментальные открытия. Так, впервые был выведен «химически чистый», изолированный ген. Впоследствии ученые открыли ферменты лигазы и рестриктазы. При помощи последних стало возможно разрезать ген на кусочки - нуклеотиды. А при помощи лигаз можно наоборот соединять, «склеивать» эти кусочки, но уже в новой комбинации, создавая, конструируя иной ген.
Значительных успехов добились ученые и в процессе «чтения» биологической информации. На протяжении многих лет расшифровкой заложенных в генах данных занимались У. Гилберт и Ф. Сенгер, американский и английский ученые.
Специалисты отмечают, что генная инженерия за весь период своего существования не оказала негативного воздействия на самих исследователей, не причинила вреда человеку и не принесла ущерба природе. Ученые отмечают, что достигнутые результаты как в процессе изучения функционирования механизмов, обеспечивающих жизнедеятельность организмов, так и в прикладной отрасли являются весьма внушительными. При этом перспективы представляются поистине фантастичными.
Несмотря на большое значение генетики и генной инженерии в сельском хозяйстве и медицине, главные ее результаты еще не достигнуты.
Перед учеными стоит достаточно много задач. Следует определить не только функции и назначение каждого гена, но и условия, при которых происходит его активация, в какие именно периоды жизни, под воздействием каких факторов, в каких именно участках организма он включается и провоцирует синтез соответствующего белка. Кроме того, немаловажно выяснить роль этого белка в жизни организма, какие реакции он запускает, выходит ли за клеточные пределы, какую несет информацию. Достаточно сложной является проблема сворачивания белков. Решение этих и многих других задач осуществляется учеными в рамках генной инженерии.
Генная инженерия - это метод биотехнологии, который занимается исследованиями по перестройке генотипов. Генотип является не просто механическая сумма генов, а сложная, сложившаяся в процессе эволюции организмов система. Генная инженерия позволяет путем операций в пробирке переносить генетическую информацию из одного организма в другой. Перенос генов дает возможность преодолевать межвидовые барьеры и передавать отдельные наследственные признаки одних организмов другим.
Носителями материальных основ генов служат хромосомы, в состав которых входят ДНК и белки. Но гены образования не химические, а функциональные. С функциональной точки зрения ДНК состоит из множества блоков, хранящих определенный объем информации - генов. В основе действия гена лежат его способность через посредство РНК определять синтез белков. В молекуле ДНК как бы записана информация, определяющая химическую структуру белковых молекул. Ген - участок молекулы ДНК, в котором находится информация о первичной структуре какого-либо одного белка (один ген - один белок). Поскольку в организмах присутствуют десятки тысяч белков, существуют и десятки тысяч генов. Совокупность всех генов клетки составляет ее геном. Все клетки организма содержат одинаковый набор генов, но в каждой из них реализуется различная часть хранимой информации. Поэтому, например, нервные клетки и по структурно-функциональным, и по биологическим особенностям отличаются от клеток печени.
Перестройка генотипов, при выполнении задач генной инженерии, представляет собой качественные изменения генов не связанные с видимыми в микроскопе изменениями строения хромосом. Изменения генов прежде всего связано с преобразованием химической структуры ДНК. Информация о структуре белка, записанная в виде последовательности нуклеотидов, реализуется в виде последовательности аминокислот в синтезируемой молекуле белка. Изменение последовательности нуклеотидов в хромосомной ДНК, выпадение одних и включение других нуклеотидов меняют состав образующихся на ДНК молекулы РНК, а это, в свою очередь, обуславливает новую последовательность аминокислот при синтезе. В результате в клетке начинает синтезироваться новый белок, что приводит к появлению у организма новых свойств. Сущность методов генной инженерии заключается в том, что в генотип организма встраиваются или исключаются из него отдельные гены или группы генов. В результате встраивания в генотип ранее отсутствующего гена можно заставить клетку синтезировать белки, которые ранее она не синтезировала.
Наиболее распространенным методом генной инженерии является метод получения рекомбинантных, т.е. содержащих чужеродный ген, плазмид. Плазмиды представляют собой кольцевые двухцепочные молекулы ДНК, состоящие из нескольких тысяч пар нуклеотидов. Этот процесс состоит из нескольких этапов.
1. Рестрикция - разрезание ДНК, например, человека на фрагменты.
2. Лигирование - фрагмент с нужным геном включают в плазмиды и сшивают их.
3. Трансформация -введение рекомбинантных плазмид в бактериальные клетки. Трансформированные бактерии при этом приобретают определенные свойства. Каждая из трансформированных бактерий размножается и образует колонию из многих тысяч потомков - клон.
4. Скрининг - отбор среди клонов трансформированных бактерий тех, которые плазмиды, несущие нужный ген человека.
Весь этот процесс называется клонированием. С помощью клонирования можно получить более миллиона копий любого фрагмента ДНК человека или другого организма. Если клонированный фрагмент кодирует белок, то экспериментально можно изучить механизм, регулирующий транскрипцию этого гена, а также наработать этот белок в нужном количестве. Кроме того, клонированный фрагмент ДНК одного организма можно ввести в клетки другого организма. Этим можно добиться, например, высокие и устойчивые урожаи благодаря введенному гену, обеспечивающему устойчивость к ряду болезней. Если ввести в генотип почвенных бактерий гены других бактерий, обладающих способностью связывать атмосферный азот, то почвенные бактерии смогут переводить этот азот в связанный азот почвы. Введя в генотип бактерии кишечной палочки ген из генотипа человека, контролирующий синтез инсулина, ученые добились получения инсулина при посредстве такой кишечной палочки. При дальнейшем развитии науки станет возможным введение в зародыш человека недостающих генов, и тем самым позволит избежать генетических болезней.
Эксперименты по клонированию животных ведутся давно. Достаточно убрать из яйцеклетки ядро, имплантировать в нее ядро другой клетки, взятой из эмбриональной ткани, и вырастить ее - либо в пробирке, либо в чреве приемной матери. Клонированная овечка Доли была создана нетрадиционным путем. Ядро из клетки вымени 6-летней взрослой овцы одной породы пересадили в безъядерное яйцо овцы другой породы. Развивающийся зародыш поместили в овцу третей породы. Так как родившаяся овечка получила все гены от первой овцы - донора, то является ее точной генетической копией. Этот эксперимент открывает массу новых возможностей для клонирования элитных пород, взамен многолетней селекции.
Ученые Техасского университета смогли продлить жизнь нескольких типов человеческих клеток. Обычно клетка умирает, пережив около 7-10 процессов деления, а они добились сто делений клетки. Старение, по мнению ученых, происходит из-за того, что клетки при каждом делении теряют теломеры, молекулярные структуры, которые располагаются на концах всех хромосом. Ученые имплантировали в клетки открытый ими ген, отвечающий за выработку теломеразы и тем самым сделали их бессмертными. Возможно это будущий путь к бессмертию.
Еще с 80-х годов появились программы по изучению генома человека. В процессе выполнения этих программ уже прочитано около 5 тысяч генов (полный геном человека содержит 50-100 тысяч). Обнаружен ряд новых генов человека. Генная инженерия приобретает все большее значение в генотерапии. Потому, что многие болезни заложены на генетическом уровне. Именно в геноме заложена предрасположенность ко многим болезням или стойкость к ним. Многие ученые считают, что в XXI веке будет функционировать геномная медицина и генная инженерия.
Сложно найти в современном мире человека, который ничего не слышал бы об успехах генной инженерии.
Сегодня она является одним из наиболее перспективных путей развития биотехнологий, совершенствования сельскохозяйственного производства, медицины и ряда других отраслей.
Что такое генная инженерия?
Как известно, наследственные признаки любого живого существа записаны в каждой клетке организма в виде совокупности генов – элементов сложных белковых молекул . Вводя в геном живого существа чужеродный ген, можно изменить свойства получаемого организма, причём в нужную сторону: сделать сельскохозяйственную культуру более устойчивой к морозу и болезням, придать растению новые свойства и т.д.
Организмы, полученные в результате такой переделки, называются генно-модифицированными, или трансгенными, а научная дисциплина, занимающаяся исследованием модификаций и разработкой трансгенных технологий – генетической или генной инженерией.
Объекты генной инженерии
Наиболее часто объектами для исследования генной инженерии становятся микроорганизмы, клетки растений и низших животных, однако ведутся исследования и на клетках млекопитающих, и даже на клетках человеческого организма. Как правило, непосредственным объектом исследования является молекула ДНК, очищенная от прочих клеточных веществ. При помощи энзимов ДНК расщепляется на отдельные отрезки, причём важно уметь распознавать и выделять нужный отрезок, переносить его при помощи энзимов и встраивать в структуру другой ДНК.
Современные методики уже позволяют достаточно свободно манипулировать отрезками генома, размножать нужный участок наследственной цепи и вставлять его на место другого нуклеотида в ДНК реципиента. Накоплен достаточно большой опыт и собрана немалая информация по закономерностям строения наследственных механизмов. Как правило, преобразованиям подвергаются сельскохозяйственные растения, что уже позволило существенно повысить результативность основных продовольственных культур.
Для чего нужна генная инженерия?
К середине ХХ века традиционные методы перестали устраивать учёных, так как это направление обладает рядом серьёзных ограничений:
- невозможно скрещивать неродственные виды живых существ;
- процесс рекомбинации генетических признаков остаётся неуправляемым, и необходимые качества у потомства появляются в результате случайных комбинаций, при этом очень большой процент потомства признаётся неудачным и отбрасывается в ходе селекции;
- точно задать нужные качества при скрещивании невозможно;
- селекционный процесс занимает годы и даже десятилетия.

Естественный механизм сохранения наследственных признаков является чрезвычайно стойким, и даже появление потомства с нужными качествами не даёт гарантии сохранения этих признаков в последующих поколениях.
Генная инженерия позволяет преодолеть все вышеперечисленные затруднения. С помощью трансгенных технологий можно создавать организмы с заданными свойствами, заменяя отдельные участки генома другими, взятыми у живых существ, принадлежащих к другим видам. При этом сроки создания новых организмов существенно сокращаются. Необязательно закреплять нужные признаки, делая их наследуемыми, так как всегда есть возможность генетически модифицировать следующие партии, поставив процесс буквально на поток.
Этапы создания трансгенного организма
- Выделение изолированного гена с нужными свойствами. Сегодня для этого существуют достаточно надёжные технологии, есть даже специально подготовленные библиотеки генов.
- Ввод гена в вектор для переноса. Для этого создаётся специальная конструкция – трансген, с одним или несколькими отрезками ДНК и регуляторными элементами, который встраивается в геном вектора и подвергается клонированию при помощи лигаз и рестриктаз. В качестве вектора обычно используются кольцеобразные бактериальные ДНК – плазмиды.
- Встраивание вектора в организм реципиента. Этот процесс скопирован с аналогичного природного процесса встраивания ДНК вируса или бактерии в клетки носителя и действует таким же образом.
- Молекулярное клонирование. При этом клетка, подвергшаяся модификации, успешно делится, производя множество новых дочерних клеток, которые содержат изменённый геном и синтезируют белковые молекулы с заданными свойствами.
- Отбор ГМО. Последний этап ничем не отличается от обычной селекционной работы.
Безопасна ли генная инженерия?
Вопрос, насколько безопасны трансгенные технологии, периодически поднимается как в научной среде, так и в СМИ, далёких от науки. Однозначного ответа на него нет до сих пор.
Во-первых, генная инженерия остаётся ещё достаточно новым направлением биотехнологий, и статистика, позволяющая делать объективные выводы об этой проблеме, пока что не успела накопиться.
Во-вторых, огромные вложения в генную инженерию со стороны транснациональных корпораций, занимающихся производством продуктов питания, могут служить дополнительной причиной отсутствия серьёзных исследований.
Впрочем, в законодательствах многих стран появились нормы, обязывающие производителей указывать наличие продуктов из ГМО на упаковке товаров пищевой группы. В любом случае, генная инженерия уже продемонстрировала высокую результативность своих технологий, а её дальнейшее развитие обещает людям ещё больше успехов и достижений.
Отправить свою хорошую работу в базу знаний просто. Используйте форму, расположенную ниже
Студенты, аспиранты, молодые ученые, использующие базу знаний в своей учебе и работе, будут вам очень благодарны.
Размещено на http://www.allbest.ru/
Генетическая инженерия, совокупность методов биохимии и молекулярной генетики, с помощью которых осуществляется направленное комбинированное генетической информации любых организмов.
Генетическая инженерия позволяет преодолевать природные межвидовые барьеры, препятствующие обмену генетической информацией между таксономически удаленными видами организмов, и создавать клетки и организмы с не существующими в природе сочетаниями генов, с заданными наследуемыми свойствами. Главным объектом генно-инженерного воздействия является носитель генетической информации - дизоксирибонуклеиновая кислота (ДНК), молекула которой обычно состоит из двух цепей. Строгая специфичность спаривания пуриновых и пиримидиновых оснований обусловливает свойство комплементарности - взаимного соответствия нуклеотидов в двух цепях. Создание новых сочетаний генов оказалось возможным благодаря принципиальному сходству строения молекул ДНК у всех видов организмов, а фактически универсальность генетического кода обеспечивает экспрессию чужеродных генов (проявление их функциональной активности) в любых видах клеток. Этому способствовало также накопление знаний в области химии нуклеиновых кислот, выявление молекулярных особенностей организации и функционирования генов (в т. ч.установление механизмов регуляции их экспрессии и возможности подчинения генов действию «чужих» регуляторных элементов), разработка методов секвенирования ДНК, открытие полимеразной цепной реакции, позволившей быстро синтезировать любой фрагмент ДНК. Важными предпосылками для появления генетической инженерии явились: открытие плазмид, способных к автономной репликации и переходу из одной бактериальной клетки в другую, и явления трансдукции - переноса некоторых генов бактериофагами, что позволило сформулировать представление о векторах: молекулах - переносчиках генов. Огромное значение в развитии методологии генетической инженерии сыграли ферменты, участвующие в преобразовании нуклеиновых кислот: рестриктазы (узнают в молекулах ДНК строго определенные последовательности - сайты - и «разрезают» двойную цепь в этих местах), ДНК-лигазы (ковалентно связывают отдельные фрагменты ДНК), обратная транскриптаза (синтезирует на матрице РНК комплементарную копию ДНК, или кДНК) и др. Только при их наличии создание искусственных структур стало технически выполнимой задачей. Ферменты используются для получения индивидуальных фрагментов ДНК (генов) и создания молекулярных гибридов - рекомбинантных ДНК (рекДНК) на основе ДНК плазмид и вирусов. Последние доставляют нужный ген в клетку хозяина, обеспечивая там его размножение (клонирование) и образование конечного продукта гена (его экспрессию).
Принципы созд ания рекомбинантных молекул ДНК
Термин «Генетическая инженерия» получил распространение после того, как в 1972 П. Бергом с сотрудниками впервые была получена рекомбинантная ДНК, представлявшая собой гибрид, в котором были соединены фрагменты ДНК бактерии кишечной палочки, ее вируса (бактериофага a) и ДНК обезьяньего вируса SV40. В 1973 С. Коэн с сотрудниками использовали плазмиду pSC101 и рестриктазу (EcoRI), которая раскрывает ее в одном месте таким образом, что на концах двухцепочечной молекулы ДНК образуются короткие комплементарные одноцепочечные «хвосты» (обычно 4 - 6 нуклеотидов). Их называли «липкими», поскольку они могут спариваться (как бы слипаться) друг с другом. Когда такую ДНК смешивали с фрагментами чужеродной ДНК, обработанной той же рестриктазой и имеющей такие же липкие концы, получались новые гибридные плазмиды, каждая из которых содержала, по крайней мере, один фрагмент чужеродной ДНК, встроенной в EcoRI-сайт плазмиды. Стало очевидным, что в такие плазмиды можно встраивать фрагменты разнообразных чужеродных ДНК, полученных как из микроорганизмов, так и из высших эукариот.
Основная современная стратегия получения рекДНК сводится к следующему:
1) В ДНК плазмиды или вируса, способных размножаться независимо от хромосомы, встраивают принадлежащие другому организму фрагменты ДНК, содержащие определенные гены или искусственно полученные последовательности нуклеотидов, представляющие интерес для исследователя;
2) Образующиеся при этом гибридные молекулы вводят в чувствительные прокариотические или эукариотические клетки, где они реплицируются (размножаются, амплифицируются) вместе со встроенными в них фрагментами ДНК;
3) Отбирают клоны клеток в виде колоний на специальных питательных средах (или вирусов в виде зон просветления - бляшек на слое сплошного роста клеток бактерий или культур тканей животных), содержащие нужные типы молекул рекДНК и подвергают их разностороннему структурно-функциональному изучению.
Для облегчения отбора клеток, в которых присутствует рекДНК, используют векторы, содержащие один и более маркеров. У плазмид, например, такими маркерами могут служить гены устойчивости к антибиотикам (отбор клетик, содержащих рекДНК, проводят по их способности расти в присутствии того или иного антибиотика). РекДНК, несущие нужные гены, отбирают и вводят в реципиентные клетки. С этого момента начинается молекулярное клонирование - получение копий рек ДНК, а следовательно, и копий целевых генов в ее составе. Только при возможности разделения всех трансфицированных или инфицированных клеток каждый клон будет представлен отдельной колонией клеток и содержать определенную рек ДНК. На заключительном этапе производится идентификация клонов, в которых заключен нужный ген. Одна основывается на том, что вставка в рек ДНК детерминирует какое-то уникальное свойство содержащей его клетки (например, продукт экспрессии встроенного гена). В опытах по молекулярному клонированию соблюдаются 2 основных принципа: ни одна из клеток, где происходит клонирование рек ДНК, не должнаполучить более одной плазмидной молекулы или вирусной частицы; последние должны быть способны к репликации.
В качестве векторных молекул в генетической инженерии используется широкий спектр плазмидных и вирусных ДНК. Наиболее популярны клонирующие векторы, содержащие несколько генетических маркеров и имеющие по одному месту действия для разных рестриктаз. Таким требованиям, например, лучше всего отвечает плазмида pBR322, которая была сконструирована из исходно существующей в природе плазмиды с помощью методов, применяемых при работе с рекДНК; она содержит гены устойчивости к ампициллину и тетрациклину, а также по одному сайту узнавания для 19 разных рестриктаз. Частным случаем клонирующих векторов являются экспрессирующие векторы, которые наряду с амплфикацией обеспечивают правильную и эффективную экспрессию чужеродных генов в реципиентных клетках. В ряде случаев молекулярные векторы могут обеспечивать интеграцию чужеродной ДНК в геном клетки или вируса (их называют интегративными векторами).
Одна из важнейших задач генетической инженерии - создание штаммов бактерий или дрожжей, линий клеток тканей животных или растений, а также трансгенных растений и животных, которые обеспечивали бы эффективную экспрессию клонируемых в них генов. Высокий уровень продукции белков достигается в том случае, если гены клонируются в многокопийных векторах, т. к. при этом целевой ген будет находиться в клетке в большом количестве. Важно, чтобы кодирующая последовательность ДНК находилась под контролем промотора, который эффективно узнается РНК-полимеразой клетки, а образующаяся мРНК была бы относительно стабильной и эффективно транслировалась. Кроме того, чужеродный белок, синтезируемый в реципиентных клетках, не должен подвергаться быстрой деградации внутриклеточными протеазами. При создании трансгенных животных и растений часто добиваются тканеспецифичной экспрессии вводимых целевых генов.
Поскольку генетический код универсален, возможность экспрессии гена определяется лишь наличием в его составе сигналов инициации и терминации транскрипции и трансляции, правильно узнаваемых хозяйской клеткой. Т. к. большинство генов высших эукариот имеет прерывистую экзон-интронную структуру, в результате транскрипции таких генов образуется матричная РНК-предшественник, из которой при последующем сплайсинге выщепляются некодирующие последовательности - интроны и образуется зрелая мРНК. Такие гены не могут экспрессироваться в клетках бактерий, где отсутствует система сплайсинга. Для того чтобы преодолеть это препятствие, на молекулах зрелой мРНК с помощью обратной транскриптазы синтезируют ДНК-копию (кДНК), к которой с помощью ДНК-полимеразы достраивается вторая цепь. Такие фрагменты ДНК, соответствующие кодирующей последовательности генов (уже не разделенной интронами), можно встраивать в подходящий молекулярный вектор.
Зная аминокислотную последовательность целевого полипептида, можно синтезировать кодирующую его нуклеотидную последовательность, получив ген-эквивалент, и встроить его в соответствующий экспрессирующий вектор. При создании гена-эквивалента обычно учитывают свойство вырожденности генетического кода (20 аминокислот кодируются 61 кодоном) и частоту встречаемости кодонов для каждой аминокислоты в тех клетках, в которые планируется вводить этот ген, т. к. состав кодонов может существенно отличаться у разных организмов. Правильно подобранные кодоны могут значительно повысить продукцию целевого белка в реципиентной клетке.
Значение генетической инженерии
Генетическая инженерия значительно расширила экспериментальные границы молекулярной биологии, поскольку стало возможным вводить в различные типы клеток чужеродную ДНК и исследовать ее функции. Это позволило выявлять общебиологические закономерности организации и выражения генетической информации в различных организмах. Данный подход открыл перспективы создания принципиально новых микробиологических продуцентов биологически активных веществ, а также животных и растений, несущих функционально активные чужеродные гены. Многие ранее недоступные биологически активные белки человека, в т. ч. интерфероны, интерлейкины, пептидные гормоны, факторы крови, стали нарабатываться в больших количествах в клетках бактерий, дрожжей или млекопитающих и широко использоваться в медицине. Более того, появилась возможность искусственно создавать гены, кодирующие химерные полипептиды, обладающие свойствами двух или более природных белков. Все это дало мощный импульс к развитию биотехнологии.
Главными объектами генетической инженерии являются бактерии Escherichia coli (кишечная палочка) и Bacillus subtilis (сенная палочка), пекарские дрожжи Saccharomices cereuisiae, различные линии клеток млекопитающих. Спектр объектов генно-инженерного воздействия постоянно расширяется. Интенсивно развиваются направления исследований по созданию трансгенных растений и животных. Методами генетической инженерии создаются новейшие поколения вакцин против различных инфекционных агентов (первая из них была создана на основе дрожжей, продуцирующих поверхностный белок вируса В человека). Большое внимание уделяется разработке клонирующих векторов на основе вирусов млекопитающих и использованию их для создания живых поливалентных вакцин для нужд ветеринарии и медицины, а также в качестве молекулярных векторов для генной терапии раковых опухолей и наследственных заболеваний. Разработан метод прямого введения в организм животных и человека рекДНК, направляющих продукцию в их клетках антигенов различных инфекционных агентов (ДНК-вакцинация). Новейшим направлением генетической инженерии является создание съедобных вакцин на основе трансгенных растений, таких как томаты, морковь, картофель, кукуруза, салат и др., продуцирующих иммуногенные белки возбудителей инфекций. генетический инженерия рекомбинантный молекула
Опасения, связанные с проведением генно-инженерных экспериментов
Вскоре после первых успешных экспериментов по получению рек ДНК группа ученых во главе с П. Бергом предложила ограничить проведение ряда генно-инженерных опытов. Эти опасения основывались на том, что свойства организмов, содержащих чужую генетическую информацию, трудно предсказать. Они могут приобрести нежелательные признаки, нарушить экологическое равновесие, привести к возникновению и распространению необычных заболеваний человека, животных, растений. Кроме того, отмечалось, что вмешательство человека в генетический аппарат живых организмов аморально и может вызвать нежелательные социальные и этические последствия. В 1975 эти проблемы обсуждались на международной конференции в Асиломаре (США). Ее участники пришли к заключению о необходимости продолжения использования методов генетической инженерии, но при обязательном соблюдении определенных правил и рекомендаций. Впоследствии эти правила, установленные в ряде стран, были существенно смягчены и свелись к приемам, обычным в микробиологических исследованиях, созданию специальных защитных устройств, препятствующих распространению биологических агентов в окружающей среде, использованию безопасных векторов и реципиентных клеток, не размножающихся в природных условиях.
Часто под генетической инженерией понимают только работу с рек ДНК, а как синонимы генетической инженерии используются термины «молекулярное клонирование», «клонирование ДНК», «клонирование генов». Однако все эти понятия отражают содержание лишь отдельных генно-инженерных операций и поэтому не эквивалентны термину «генетическая инженерия». В России как синоним генетической инженерии широко используется термин «генная инженерия». Однако смысловое содержание этих терминов различно: генетическая инженерия ставит целью создание организмов с новой генетической программой, в то время как термин «генная инженерия» поясняет, как это делается - путем манипуляции с генами.
Размещено на Allbest.ru
Подобные документы
Генная инженерия как раздел молекулярной генетики, связанный с целенаправленным созданием новых комбинаций генетического материала. История ее возникновения и развития, этапы генного синтеза. Безопасна ли генная модификация? Примеры ее применения.
реферат , добавлен 23.11.2009
Понятие и основные методы генной инженерии. Методика выделения ДНК на примере ДНК плазмид. Принципы действия системы рестрикции-модификации. Перенос и обнаружение клонируемых генов в клетках. Конструирование и введение в клетки рекомбинантных молекул ДНК.
реферат , добавлен 23.01.2010
Исследование сущности и предназначения генной инженерии - метода биотехнологии, который занимается исследованиями по перестройке генотипов. Метод получения рекомбинантных, то есть содержащих чужеродный ген, плазмид - кольцевых двухцепочных молекул ДНК.
презентация , добавлен 19.02.2012
Суть и задачи генной инженерии, история ее развития. Цели создания генетически модифицированных организмов. Химическое загрязнение как следствие ГМО. Получение человеческого инсулина как важнейшее достижение в сфере генно-модифицированных организмов.
реферат , добавлен 18.04.2013
Использование генной инженерии как инструмента биотехнологии с целью управления наследственностью живых организмов. Особенности основных методов и достижений генной инженерии в медицине и сельском хозяйстве, связанные с ней опасности и перспективы.
доклад , добавлен 10.05.2011
Генная инженерия как метод биотехнологии, который занимается исследованиями по перестройке генотипов. Этапы процесса получения рекомбинантных плазмид. Конструирование клеток нового типа на основе их культивирования, гибридизации и реконструкции.
презентация , добавлен 20.11.2011
Генная инженерия: история возникновения, общая характеристика, преимущества и недостатки. Знакомство с новейшими методами генной инженерии, их использование в медицине. Разработка генной инженерии в области животноводства и птицеводства. Опыты на крысах.
курсовая работа , добавлен 11.07.2012
Генетическая инженерия - инструмент биотехнологии для получения рекомбинантных РНК и ДНК, осуществления манипуляций с генами и белковыми продуктами, введения их в другие организмы. Современное состояние науки о наследственности и хромосомных болезнях.
реферат , добавлен 23.06.2009
Возникновение биотехнологии. Основные направления биотехнологии. Биоэнергетика как раздел биотехнологии. Практические достижения биотехнологии. История генетической инженерии. Цели, методы и ферменты генной инженерии. Достижения генетической инженерии.
реферат , добавлен 23.07.2008
Основы и техника клонирования ДНК. Этапы генной инженерии бактерий. Развитие генетической инженерии растений. Генетическая трансформация и улучшение растений с помощью агробактерий, источники генов. Безопасность генетически модифицированных растений.




